Signal transfer towards expression of alkaloid biosynthesis
Selective induction of phytoalexin biosynthesis in Eschscholzia cells - a story of images
The production of secondary metabolites in response to microbial pathogens or herbivores is a very effective element in the chemical arsenal of plant defense mechanisms. Secondary products with antimicrobial or anti-herbivorous effects are a main determinant in the evolution of secondary metabolism.
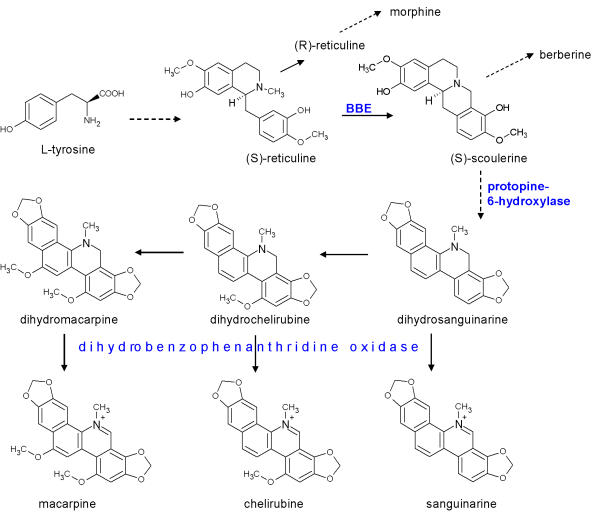
Benzophenanthridine alkaloids, notably sanguinarine, are among the strongest antimicrobial agents produced by plants. They are built from tyrosine at the benzylisoquinoline pathway, which also branches to morphine and berberine. The enzymes of this biosynthesis are all known, some of them are cloned.
In contrast to morphine, benzophenanthridines can be produced in suspensions of cultured cells. Cell suspensions of Eschscholzia californica respond to a number of stressors with the overproduction of benzophenanthridine alkaloids. They are used as a model system to investigate the expression of secondary metabolism by external stimuli.

|
Eschscholzia californica is a medicinal plant used since ancient times. In the root, it produces a great diversity of isoquinoline alkaloids, among them benzophenanthridines as sanguinarine, chelerythrine and macarpine.
|
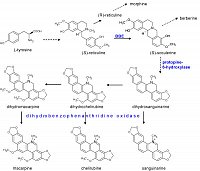
according to Haider et al. 1997
|

|
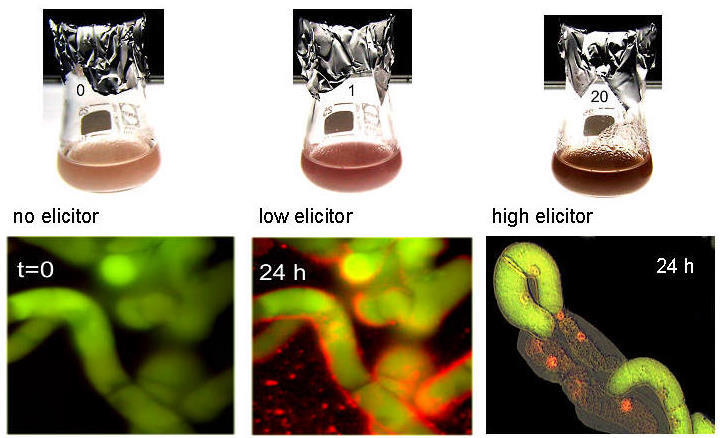
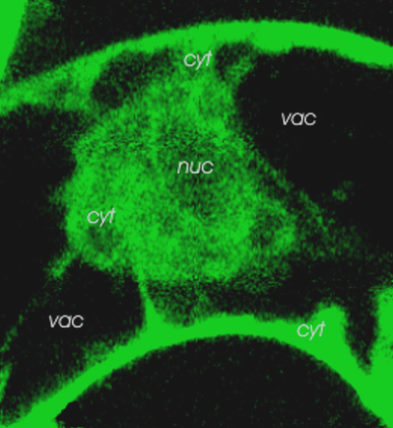
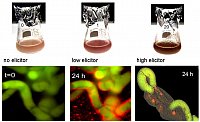
left: Callus of Eschscholzia californica meets hyphae of Penicillium cyclopium: benzophenanthridines (red color) are produced at the contact sites.
right: cell suspensions loaded with viability marker (green fluorescing).
low elicitor concentrations trigger alkaloid biosynthesis followed by excretion of the benzophenanthridines, cells remain intact.
high elicitor concentrations trigger alkaloid production plus hypersensitive response (medium alkalinization, browning, oxidative burst, cell death). (Roos et al. J.Plant Physiol. 2006)
|
|
Next images refer to the signal chain triggered by low elicitor concentrations leading to alkaloid production without cell death.
|
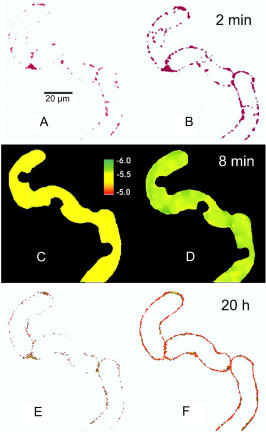
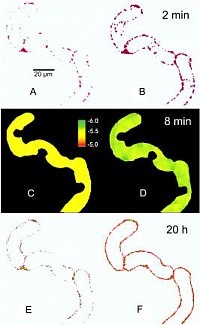
Two early signal events and the cellular response to elicitor treatment occur in the same cell:
- activation of phospholipase A2 in the plasma
membrane
(top: hydrolysis of a fluorogenic substrate
of PLA2)
- efflux of protons from the vacuole
(middle: confocal pH maps of the vacuole)
- production and excretion of benzophen-
anthridine alkaloids
(bottom: alkaloid fluorescence in the cell wall)
(Confocal microscopy, Viehweger et al., Plant Cell 2006)
|
|
|
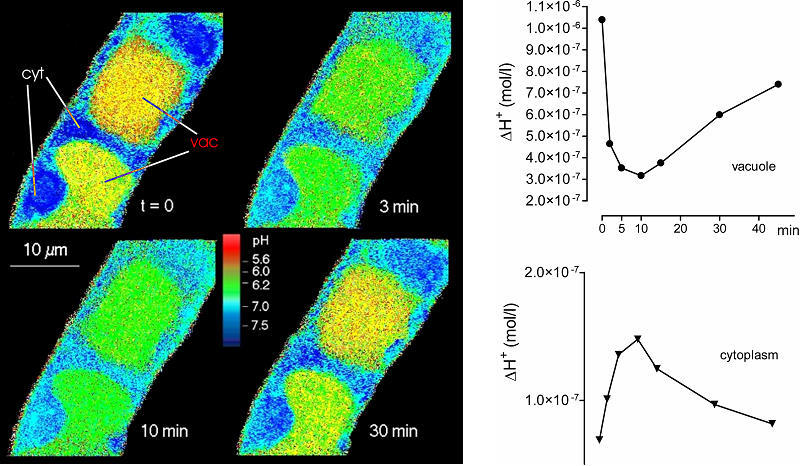
The pH shift observed after elicitor contact is brought about by a transient efflux of vacuolar protons as shown by confocal pH topography of intact cells. The image shows a pH map obtained by confocal pH topography of intact Eschscholzia cells, during perfusion with culture liquid. Elicitor was added at t = 1 min.
Trace, top: decline of H+ in the vacuole
Trace, bottom: increase of H+ in the cytoplasm
|

|
|
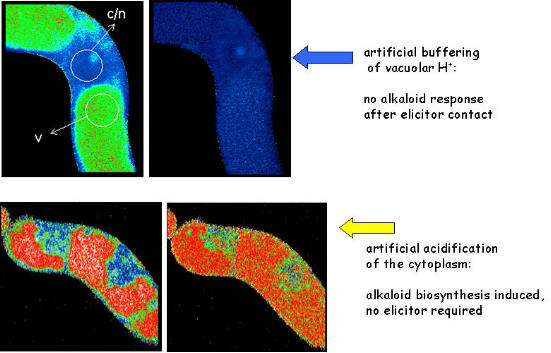
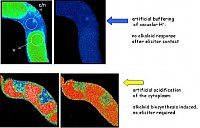
The efflux of vacuolar protons into the cytoplasm is essential for the signal transfer:
- after setting the vaculoar pH to 7.4 (shown in upper row) elicitor signaling is blocked
- experimentally triggered acidification of the cytoplasm (shown in bottom row)
induces alkaloid formation in the absence of elicitor.
(Roos et al, Plant Phys.1998, Viehweger et al. Plant Cell 2002)
|
|
|
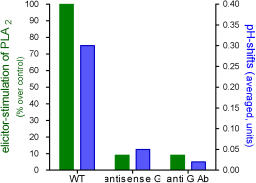
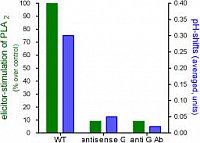
The activation of PLA2 and the pH shift depend on
a functional Galpha protein:
cell lines of low Galpha content (antisense
transformation), or cell lines expressing an
anti-Galpha antibody (scFv type) against Galpha
are deficient in both signal events.
(Viehweger et al. Plant Cell, 2006)
|
|
The activation of PLA2 and the efflux of vacuolar protons are connected via lysophosphatidylcholine (LPC), that has been identified as a new signal molecule: LPC is produced by PLA2 at the plasma membrane and traffics to the tonoplast where it activates Na+/H+ antiporters. (Viehweger et al. Plant Cell, 2006)
|
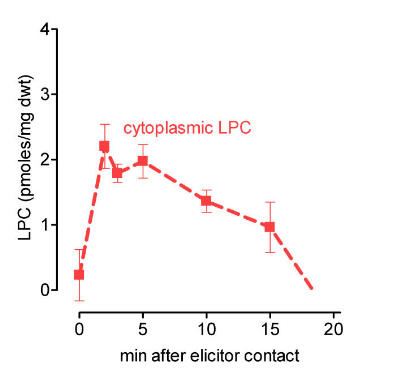
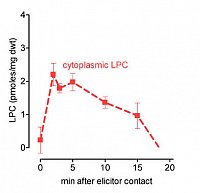
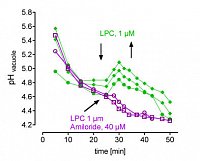
left: Cytoplasmic LPC peaks after elicitor contact, indicating activation of phospholipase A2. LPC was extracted from intact cells. It is confined to the cytoplasm as seen below.
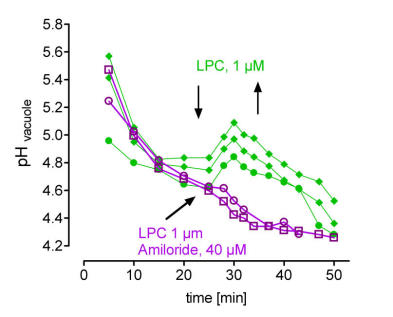
right: in situ vacuoles react to LPC with a transient loss of protons,
this response is blocked by amiloride, a specific inhibitor of Na+/H+ antiporters. Each trace represents the average pH of the central vacuole in situ, during perfusion with culture liquid. Arrows indicate addition and wirthdrawal of LPC +- amiloride (16:0).
(Viehweger et al. Plant Cell 2002).
|
|
|
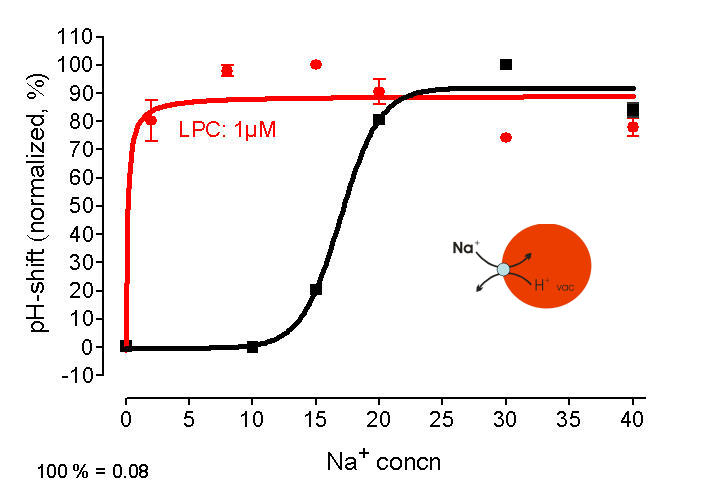
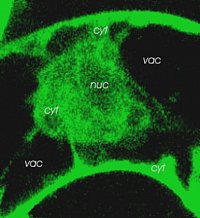
left: LPC is highly mobile and can rapidly travel from the site of genertaion (plasmalemma) to the tonoplast. Image shows the distribution of fluorescent labelled LPC as reached within 2 min.
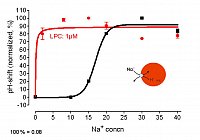
right: Na+/H+- antiport measured at vacuoles in situ - efflux of protons triggered by cytoplasmic Na+ black trace: in absence of LPC, > 10 mM extravacuolar Na+ is required red trace: in presence of LPC, 1 mM Na+ is sufficient to cause a similar efflux of H+
Conclusion: LPC increases the Na+ - sensitivity of Na+/H+- antiporter(s)
(Viehweger et al. Plant Cell 2006).
|
|
|
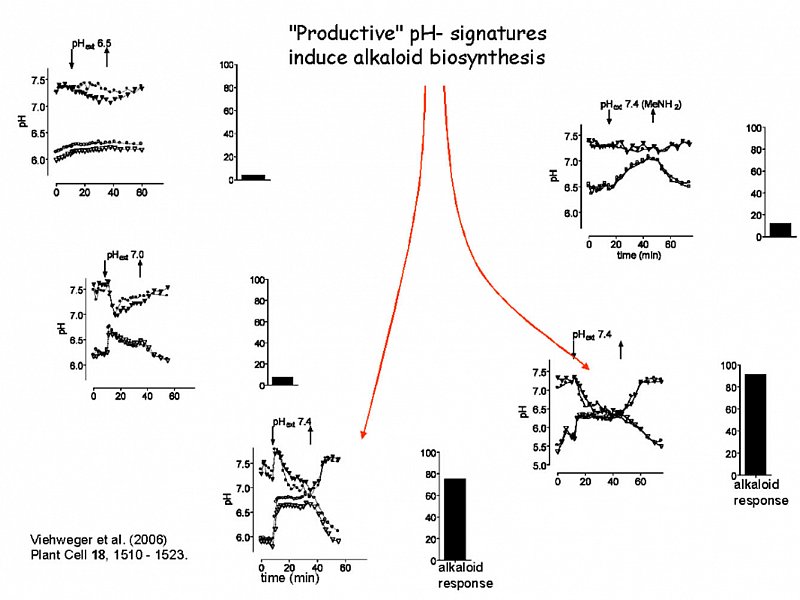
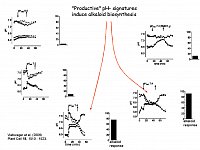
Distinct artificial pH signatures induce enzymes of alkaloid biosynthesis.
The pH traces shown here are triggered artificially by controlled perfusion of cell suspensions with strongly buffered external media of different pH. Upper traces show the average cytoplasmic pH of two typical cells, the lower traces show vacuolar pH of the same cell.
Columns indicate the increase of alkaloid biosynthesis caused by either treatment, which was reflected by the expression (RT-PCR) of the biosynthetic berberine bridge enzyme (Viehweger et al. Plant Cell 2006).
|
|
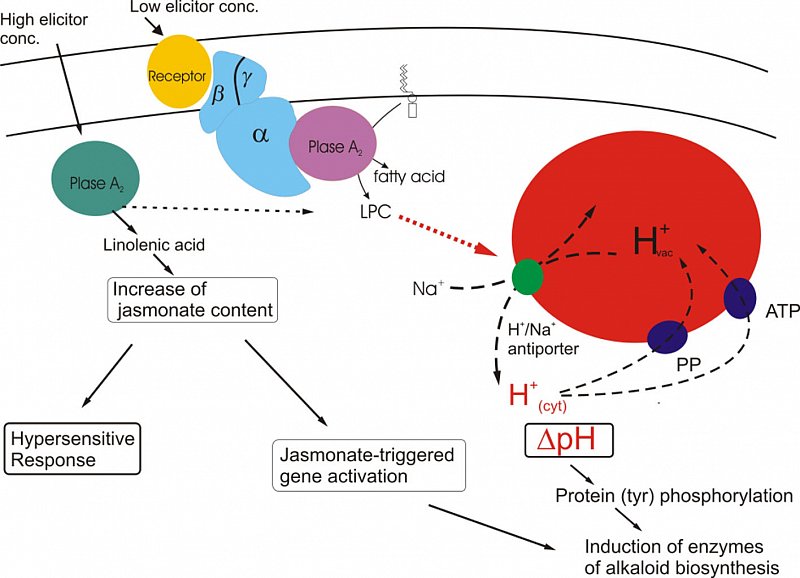
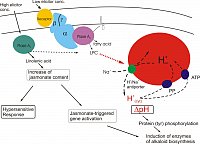
Actual summary of signaling sequences leading to the expression of alkaloid biosynthesis (Roos et al. J. Plant Physiol.2006).
Two basically different signal paths converge in the induction of biosynthetic enzymes:
low elicitor concentrations initiate a Galpha-dependent pathway that essentially includes transient pH signatures in the cytoplasm.
high elicitor concentrations signal via a peak of jasmonate, a known inducer of alkaloid biosynthesis and of the hypersensitive response. This path does not include cytoplasmic acidification or efflux of vacuolar protons (Färber et al. Phytochem. 2003).
|
Zum Seitenanfang